O sistema imunológico foi concebido para defender o corpo contra invasores estranhos ou perigosos. Tais invasores incluem
Micro-organismos (comumente chamados germes, como bactérias, vírus e fungos)
Parasitas (como vermes)
Células cancerígenas
Órgãos e tecidos transplantados
Para defender o organismo contra esses invasores, o sistema imunológico deve ter a capacidade de distinguir entre
O que pertence ao organismo (o próprio corpo)
O que não pertence (não próprio do corpo ou estranho)
Antígenos são qualquer substância que o sistema imunológico consiga reconhecer e assim estimular uma resposta imunológica. Se os antígenos forem percebidos como perigosos (por exemplo, se puderem causar uma doença) eles podem estimular uma resposta imunológica no organismo. Os antígenos podem estar em bactérias, vírus, outros micro-organismos, parasitas ou células cancerígenas. Os antígenos podem ainda existir de forma autônoma, como o pólen ou as moléculas de alimentos, por exemplo.
Uma resposta imunológica normal consiste do seguinte:
Reconhecer um antígeno estranho potencialmente nocivo
Ativar e mobilizar forças para se defender contra o antígeno
Atacar o antígeno
Controlar e finalizar o ataque
Se o sistema imunológico tiver uma disfunção e confundir o próprio corpo com algo estranho, ele pode atacar os tecidos do próprio organismo, causando uma doença autoimune, como artrite reumatoide, tireoidite de Hashimoto ou lúpus eritematoso sistêmico (lúpus).
Os distúrbios do sistema imunológico ocorrem quando
O organismo gera uma resposta imunológica contra si mesmo (uma doença autoimune).
O organismo não consegue gerar respostas imunológicas apropriadas contra os micro-organismos que o invadem (uma doença decorrente de imunodeficiência).
O corpo gera uma resposta imunológica excessiva a antígenos muitas vezes inofensivos e prejudica tecidos normais (uma reação alérgica).
Componentes do sistema imunológico
O sistema imunológico tem muitos componentes:
Anticorpos (imunoglobulinas) são proteínas produzidas por glóbulos brancos chamados células B que se ligam fortemente ao antígeno de um invasor, marcando o invasor para ser atacado ou neutralizando-o diretamente. O corpo produz milhares de anticorpos diferentes. Cada anticorpo é específico para um determinado antígeno.
Antígenos são qualquer substância que o sistema imunológico consiga reconhecer e assim estimular uma resposta imunológica.
Células B (linfócitos B) são glóbulos brancos que produzem anticorpos específicos para o antígeno que estimulou sua produção.
Basófilos são glóbulos brancos que liberam histamina (substância presente nas reações alérgicas) e que produzem substâncias que atraem outros glóbulos brancos (os neutrófilos e os eosinófilos) ao foco do problema.
Células são a menor unidade de um organismo vivo, composta por um núcleo e um citoplasma e envolta por uma membrana.
Quimiotaxia é o processo no qual uma substância química atrai células para um local específico.
O sistema de complemento consiste de um grupo de proteínas que está envolvido em uma série de reações (denominada de cascata do complemento) e que foi concebido para defender o organismo – por exemplo, destruindo bactérias e outras células estranhas, tornando as células estranhas mais fáceis de serem identificadas e ingeridas pelos macrófagos e atraindo os macrófagos e os neutrófilos para o foco do problema.
Citocinas são várias proteínas diferentes secretadas pelas células imunológicas e por outras células e que atuam como mensageiras do sistema imunológico para ajudar a regular uma resposta imunológica.
Células dendríticas são derivadas de glóbulos brancos. Elas residem nos tecidos e ajudam as células T a reconhecer antígenos estranhos.
Eosinófilos são glóbulos brancos que matam bactérias e outras células estranhas grandes demais para serem ingeridas e podem ajudar a imobilizar e matar parasitas e ajudar a destruir células cancerígenas. Eosinófilos também participam das reações alérgicas.
Células T helper são glóbulos brancos que ajudam as células B a produzir anticorpos contra antígenos estranhos, ajudam na ativação das células T killer e estimulam os macrófagos, possibilitando que eles ingiram células infectadas ou anormais com maior eficiência.
Histocompatibilidade (literalmente, compatibilidade de tecidos) é determinada por antígenos de leucócitos humanos (moléculas de autoidentificação). Histocompatibilidade é usada para determinar se um tecido ou órgão transplantado será aceito pelo receptor.
Antígenos de leucócitos humanos (HLA) são um grupo de moléculas de identificação localizadas na superfície de todas as células em uma combinação única para cada indivíduo, desta forma permitindo ao organismo distinguir o que é próprio do que não é próprio do corpo. Este grupo de moléculas de identificação é também chamado o complexo principal de histocompatibilidade.
Um complexo imunológico é um anticorpo ligado a um antígeno.
Uma resposta imunológica é a reação do sistema imunológico a um antígeno.
Imunoglobulina é outro nome para anticorpo.
Interleucina é um tipo de mensageiro (citocina) produzido por alguns glóbulos brancos para afetar outros glóbulos brancos.
Células T killer (citotóxicas) são células T que se ligam a células infectadas e células cancerígenas e as matam.
Leucócito é outro nome para glóbulo branco, como um monócito, neutrófilo, eosinófilo, basófilo ou linfócito (uma célula B ou célula T).
O sistema linfático é uma rede de linfonodos conectados por vasos linfáticos, que ajudam o corpo a transportar micro-organismos e a filtrar e destruir células mortas ou danificadas. Respostas imunes adquiridas são iniciadas nos linfonodos.
Linfócitos são os glóbulos brancos responsáveis pela imunidade adquirida (específica), inclusive a produção de anticorpos (pelas células B) e a distinção entre o que é próprio do que não é próprio do corpo (pelas células T), e por matar células infectadas e as células cancerígenas (pelas células T killer).
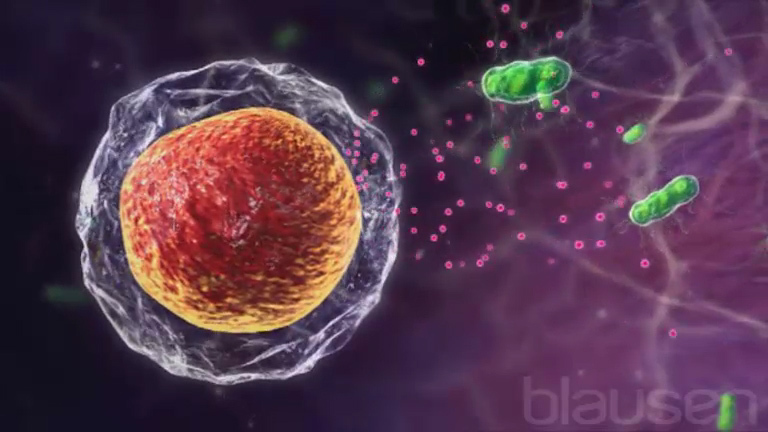
Macrófagos são células grandes que se desenvolvem a partir de glóbulos brancos denominados monócitos. Eles ingerem bactérias e outras células estranhas e ajudam as células T helper a identificar micro-organismos e outras substâncias estranhas. Macrófagos estão normalmente presentes nos pulmões, pele, fígado e outros tecidos.
Complexo principal de histocompatibilidade (MHC) é um sinônimo para antígenos de leucócitos humanos.
Mastócitos são células nos tecidos que liberam histamina e outras substâncias envolvidas nas reações inflamatórias e alérgicas.
Uma molécula é um grupo de átomos combinados quimicamente que formam uma substância única.
Células natural killer são um tipo de glóbulos brancos que reconhecem e matam células anormais, como algumas células infectadas e células cancerígenas, sem ter que saber antes que as células são anormais.
Neutrófilos são glóbulos brancos que ingerem e matam bactérias e outras células estranhas.
Fagócitos são um tipo de célula que ingere e mata ou destrói micro-organismos invasores, outras células e fragmentos de células. Fagócitos incluem neutrófilos e macrófagos.
Fagocitose é o processo em que uma célula traga e ingere um micro-organismo invasor, outra célula ou um fragmento de célula.
Um receptor é uma molécula localizada na superfície de uma célula ou dentro da célula capaz de identificar moléculas específicas, que se encaixam exatamente, como chave e fechadura.
Células T regulatórias (supressoras) são glóbulos brancos que ajudam a finalizar uma resposta imunológica.
Células T (linfócitos T) são glóbulos brancos que estão envolvidos na imunidade adquirida. Existem três tipos: helper, killer (citotóxicas) e regulatórias.
Glóbulos brancos (leucócitos) existem como vários tipos diferentes, como monócitos, neutrófilos, eosinófilos, basófilos e linfócitos (células B e células T), cada um com funções diferentes no sistema imunológico.
Linhas de defesa
O organismo tem uma série de defesas. As defesas incluem
Barreiras físicas
Glóbulos brancos
Moléculas, como anticorpos e proteínas do complemento
Órgãos linfoides
Barreiras físicas
A primeira linha de defesa contra os invasores é constituída por barreiras mecânicas ou físicas:
A pele
A córnea dos olhos
As membranas que revestem os aparelhos respiratório, digestivo, urinário e reprodutor
Enquanto estas barreiras permanecem intactas, muitos invasores não conseguem penetrar o organismo. Quando uma barreira se rompe (por exemplo, quando uma queimadura extensa lesiona a pele), o risco de infecção aumenta.
Além disso, as barreiras encontram-se defendidas por secreções que contêm enzimas que podem destruir as bactérias. O suor, as lágrimas nos olhos, as secreções no aparelho respiratório e digestivo e as secreções na vagina são exemplos de barreiras.
Glóbulos brancos
A linha de defesa seguinte envolve glóbulos brancos (leucócitos) que se deslocam através da corrente sanguínea e penetram nos tecidos para detectar e atacar micro-organismos e outros invasores.
Esta defesa consiste de duas partes:
Imunidade inata
Imunidade adquirida
Imunidade inata (natural): Inata significa algo com a qual a pessoa nasce. Então, imunidade inata não requer encontros anteriores com um micro‑organismo ou outros invasores para funcionar com eficiência. Responde imediatamente aos invasores, sem precisar aprender a reconhecê-los. Estão presentes diversos tipos de glóbulos brancos:
Os fagócitos ingerem os invasores. Fagócitos incluem macrófagos, neutrófilos, monócitos e células dendríticas.
Células natural killer são formadas prontas para reconhecer e matar as células cancerígenas e células infectadas por certos vírus.
Alguns glóbulos brancos (como basófilos e eosinófilos) liberam substâncias envolvidas em inflamações, como citocinas e em reações alérgicas, como a histamina. Algumas destas células conseguem destruir invasores diretamente.
Imunidade adquirida (adaptativa ou específica): Na imunidade adquirida, os glóbulos brancos chamados linfócitos (células B e células T) se deparam com um invasor, aprendem a atacá-lo e lembram do invasor específico de modo que possam atacá-lo de forma ainda mais eficaz no próximo encontro. A imunidade adquirida demora a desenvolver depois de um encontro inicial com um novo invasor pois os linfócitos têm que se adaptar. Mas, depois deste tempo, a resposta é rápida. As células B e as células T trabalham juntas para destruir os invasores. Para serem capazes de reconhecer invasores, as células T precisam de ajuda das células chamadas células apresentadoras de antígenos (como células dendríticas – veja a figura Como uma célula T reconhece os antígenos). Estas células ingerem os invasores ou decompõem-nos em fragmentos.
Moléculas
A imunidade inata e a imunidade adquirida interagem, influenciando-se de forma recíproca e direta ou através de moléculas que atraem ou ativam outras células do sistema imunológico, como parte da fase de mobilização das defesas. Essas moléculas incluem
Citocinas (que são as mensageiras do sistema imunológico)
Proteínas do complemento (que formam o sistema de complemento)
Estas substâncias não estão contidas nas células, encontrando-se dissolvidas em um fluido do organismo, o plasma (a parte líquida do sangue).
Algumas destas moléculas, inclusive algumas citocinas, estimulam a inflamação.
A inflamação ocorre, pois estas moléculas atraem as células do sistema imunológico ao tecido afetado. De forma a ajudar estas células a chegarem ao tecido, o organismo envia mais sangue ao tecido. Para mais sangue chegar ao tecido, os vasos sanguíneos dilatam-se e tornam-se mais porosos, permitindo que mais fluidos e células saiam dos vasos sanguíneos e penetrem o tecido. Desta forma, a inflamação tende a causar vermelhidão, calor e inchaço. O objetivo da inflamação é conter a infecção de forma que ela não se propague. As outras substâncias produzidas pelo sistema imunológico ajudam a resolver a inflamação e os tecidos danificados são curados. Apesar de a inflamação ser incômoda, é uma indicação de que o sistema imunológico está fazendo seu trabalho. Entretanto, inflamações excessivas ou de longa duração (crônicas) podem ser prejudiciais.
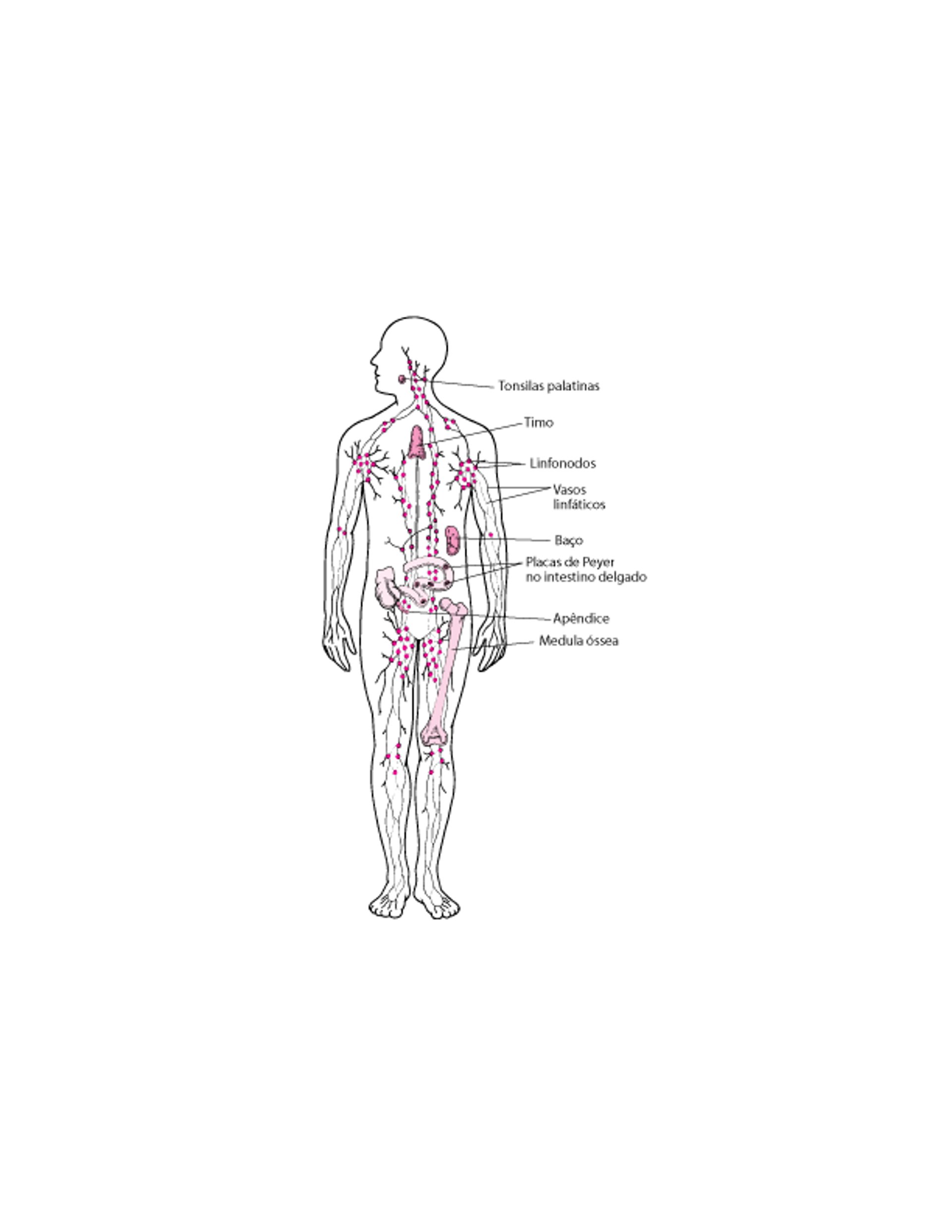
Órgãos linfoides
O sistema imunológico é constituído por vários órgãos, além de células dispersas por todo o corpo. Esses órgãos classificam-se em órgãos linfoides primários ou secundários.
Nos órgãos linfoides primários os glóbulos brancos são produzidos e/ou multiplicados:
A medula óssea produz todos os diferentes tipos de glóbulos brancos, inclusive neutrófilos, eosinófilos, basófilos, monócitos, células B e células que se tornam as células T (células T precursoras).
No timo, as células T se multiplicam e são treinadas para reconhecer antígenos estranhos e ignorar os antígenos próprios do organismo. As células T são essenciais na imunidade adquirida.
Os glóbulos brancos, especialmente da medula óssea, são mobilizados quando é preciso defender o organismo. Entram, de imediato, na corrente sanguínea e deslocam-se até à parte do corpo onde sua presença é necessária.
Os órgãos linfoides secundários incluem o
Baço
Linfonodos
Amigdalas
Apêndice
Placas de Peyer no intestino delgado
Esses órgãos retêm os micro-organismos e outras substâncias estranhas e proporcionam um lugar onde as células maduras do sistema imunológico se acumulam, interagem umas com as outras e com as substâncias estranhas, gerando uma resposta imunológica específica.
Os linfonodos encontram-se dispostos de forma estratégica no organismo, encontrando-se ligados entre si por uma extensa rede de vasos linfáticos – o sistema linfático. O sistema linfático transporta micro-organismos, outras substâncias estranhas, células cancerígenas e células mortas ou lesionadas dos tecidos aos linfonodos, onde estas substâncias e células são filtradas ou destruídas. Uma vez filtrada, a linfa é devolvida à corrente sanguínea.
Os linfonodos constituem um dos primeiros órgãos aos quais se podem propagar as células cancerígenas. Por este motivo, o médico costuma examinar os linfonodos para determinar se um câncer se propagou ou não. Células cancerígenas em um nódulo linfático podem causar inchaço no nódulo. Os linfonodos também podem inchar após uma infecção, pois as respostas imunológicas adquiridas contra as infecções são geradas nos linfonodos. Os linfonodos podem, por vezes, inchar, pois as bactérias levadas aos linfonodos não são eliminadas e causam uma infecção no linfonodo (linfadenite).
Sistema linfático: Ajuda a proteger contra as infecções
O sistema linfático constitui uma parte vital do sistema imunológico, juntamente com o timo, a medula óssea, o baço, as amígdalas, o apêndice e as placas de Peyer no intestino delgado. O sistema linfático consiste de uma rede de gânglios linfáticos que se encontram ligados pelos vasos linfáticos. Este sistema transporta a linfa por todo o organismo. A linfa é composta por um fluido que penetra os tecidos do organismo através das paredes finas dos capilares. Este fluido contém oxigênio, proteínas, e outros nutrientes que nutrem os tecidos. Uma parte deste fluido volta a entrar nos capilares e outra parte entra nos vasos linfáticos (tornando-se a linfa). Pequenos vasos linfáticos se conectam a vasos maiores acabando por formar o canal torácico. O canal torácico é o maior vaso linfático. Junta-se à veia subclávia e assim devolve a linfa na corrente sanguínea. A linfa também transporta, para eliminação, substâncias estranhas (como bactérias), células cancerígenas e células mortas ou danificadas que podem estar presentes nos tecidos dentro dos vasos linfáticos e linfonodos. A linfa contém muitos glóbulos brancos. Todas as substâncias transportadas pela linfa passam por pelo menos um gânglio linfático, onde as substâncias estranhas podem ser filtradas e destruídas antes dos fluidos serem devolvidos na corrente sanguínea. Nos linfonodos, os glóbulos brancos podem se reunir, interagir uns com os outros e com os antígenos e gerar respostas imunológicas às substâncias estranhas. Os linfonodos contêm uma rede de tecidos repletos de células B, células T, células dendríticas e macrófagos. Os micro-organismos prejudiciais são filtrados por esta rede e, então, identificados e atacados pelas células B e pelas células T. Os linfonodos costumam agrupar-se em zonas em que os vasos linfáticos se ramificam, como o pescoço, as axilas e as virilhas. |

Você sabia que...
|
Plano de ação
Uma resposta imunológica bem-sucedida aos invasores exige
Reconhecimento
Ativação e mobilização
Regulação
Resolução
Reconhecimento
Para poder destruir os invasores, o sistema imunológico deve reconhecê-los primeiro. Ou seja, o sistema imunológico deve ser capaz de distinguir o que não é próprio do corpo (estranho) do próprio do corpo. O sistema imunológico é capaz de fazer essa distinção, visto que todas as células têm moléculas de identificação (antígenos) na sua superfície. Os micro-organismos são reconhecidos porque possuem na sua superfície moléculas específicas que os identificam como estranhos.
Nas pessoas, as principais moléculas de autoidentificação denominam-se
Antígenos de leucócitos humanos (HLA) ou complexo principal de histocompatibilidade (MHC).
As moléculas HLA são denominadas antígenos, porque se transplantadas, como em um rim ou enxertos de pele, elas podem dar origem a uma resposta imunológica em outro indivíduo (geralmente estas moléculas não provocam uma resposta imunológica no indivíduo que as possui). Cada indivíduo tem uma combinação quase única de HLAs. O sistema imunológico de cada indivíduo reconhece esta combinação única como própria. Quando uma célula apresenta na sua superfície moléculas que não são iguais às das células próprias do organismo, esta é identificada como estranha. O sistema imunológico ataca imediatamente esta célula. Essa célula pode ser uma célula de um tecido transplantado ou uma das células do corpo que foi infectada por um micro-organismo invasor ou foi alterada por câncer. (são as moléculas HLA que o médico tenta compatibilizar quando um indivíduo necessita de um transplante de órgão).
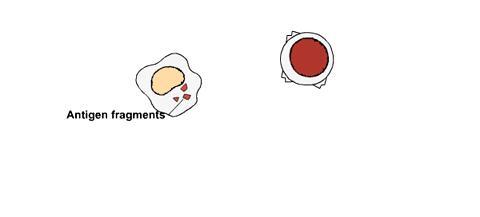
Alguns glóbulos brancos, células B (linfócitos B), conseguem reconhecer os invasores diretamente. Já outros, as células T (linfócitos T), precisam da ajuda de outras células denominadas células apresentadoras de antígenos:
Estas células ingerem os invasores e os decompõem em fragmentos.
As células apresentadoras de antígenos então combinam os fragmentos dos antígenos do invasor com as moléculas HLA da célula.
A combinação dos fragmentos de antígenos e as moléculas HLA vai para a superfície da célula.
Uma célula T com um receptor pareado na sua superfície é capaz de se fixar a parte da molécula HLA apresentadora de antígeno, como o encaixe de uma chave na fechadura correspondente.
A célula T é, por conseguinte, ativada e começa a combater os invasores que contêm aquele antígeno.
Como uma célula T reconhece antígenos
As células T fazem parte do sistema de vigilância imunológica. Elas se deslocam pela corrente sanguínea e pelo sistema linfático. Quando atingem os linfonodos ou outro órgão linfoide secundário, elas procuram substâncias estranhas (antígenos) no organismo. No entanto, antes de poderem reconhecer totalmente e responder ao antígeno estranho, o antígeno deve ser processado e apresentado à célula T por outro glóbulo branco, denominado de célula apresentadora de antígeno. As células apresentadoras de antígenos consistem em células dendríticas (que são as mais eficazes), macrófagos e células B. |

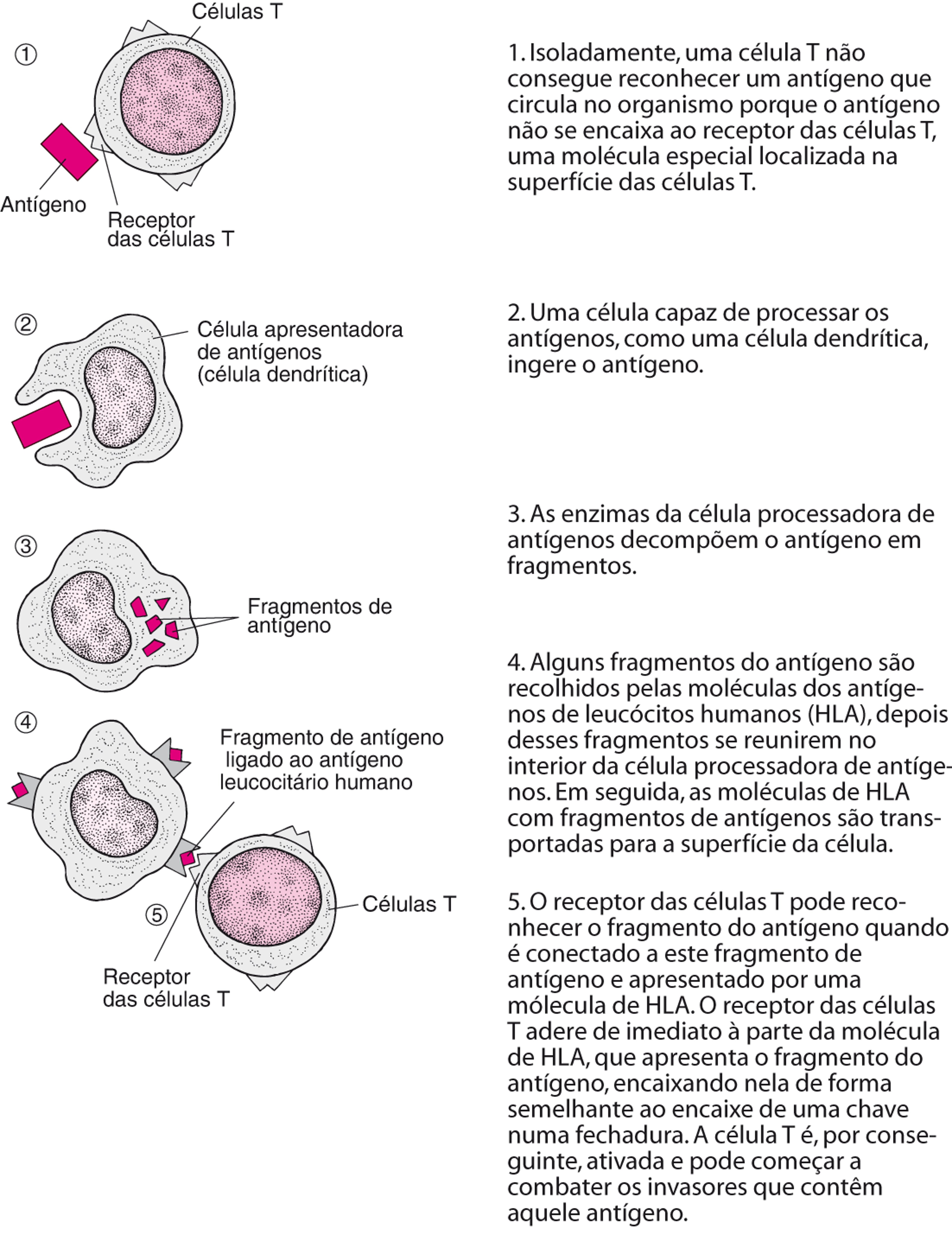
Ativação e mobilização
Os glóbulos brancos são ativados quando reconhecem os invasores. Por exemplo, quando uma célula apresentadora de antígeno apresenta fragmentos de antígenos ligados ao HLA a uma célula T, a célula T adere aos fragmentos e é ativada. As células B podem ser ativadas pelos invasores. Uma vez ativados, os glóbulos brancos tragam ou matam o invasor, ou executam ambas ações. Normalmente, mais de um tipo de glóbulos brancos é necessário para matar um invasor.
Células imunológicas, como os macrófagos ou células T ativadas, liberam substâncias que atraem outras células imunológicas ao foco do problema, mobilizando, assim, as defesas. O invasor pode também liberar substâncias que atraem as células imunológicas.
Regulação
A resposta imunológica deve ser regulada de forma a evitar um dano maior ao organismo, como ocorre nas doenças autoimunes. As células T reguladoras (supressoras) ajudam a controlar a resposta ao produzir citocinas (mensageiros químicos do sistema imunológico) que inibem as respostas imunológicas. Estas células evitam que a resposta imunológica continue indefinidamente.
Resolução
A resolução implica restringir o invasor e eliminá-lo do organismo. Após o invasor ser eliminado, a maioria dos glóbulos brancos se autodestroem e são ingeridos. Os que são poupados são denominados como células de memória. O organismo retém as células de memória, que fazem parte da imunidade adquirida, de forma a lembrar invasores específicos e poder dar uma resposta de forma mais vigorosa no próximo encontro.